|
Long lived mutants implications |
|||
|
|
|
|
|
|
|
|
||
|
|
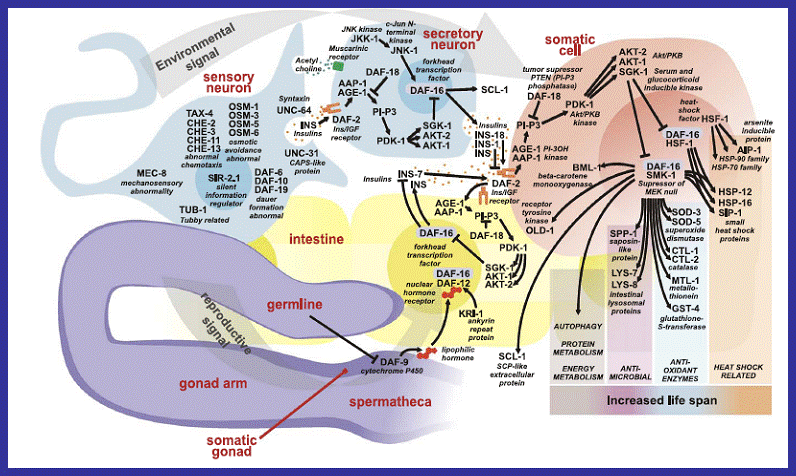
Figure
14 :
A diagram showing the complex interactions that ultimately affect lifespan in
C.elegans because of changes in
DAF-16 regulation. This image demonstrates the current knowledge about the
complexity of interactions within even a relatively simple organism. It also
shows the effect of environmental signals on the overall pathways. The red
arrows were inserted to highlight the position of the age-1 locus in the pathways (Braeckman and
Vanfleteren, 2006). |
||
|
|
|
||
|
|
Fitness vs. Lifespan in nematodes It turns out that the presence of long-lived
mutants does not violate current or historical theories about ageing
evolution. In fact the presence of these genes helps to validate the
pleiotropy theory that genes beneficial to fitness early in life will be
selected for even if they act to reduce longevity. A study by Walker et al,
confirmed that fitness does decrease in nematodes carrying the long-life
age-1 gene but only after involving other factors. As previously described,
at 20oC age-1 mutants and wild type nematode do not differ in
observerable characters except for lifespan. Researchers cultured
self-fertilizing populations of mutants and wild-type C. elegans on agar plates at 20oC. Throughout the
experiment they determined the relative allele frequencies at the age-1 locus by increasing the
temperature to 27oC where mutant nematodes develop into dauer
larvae and wild-types remain adults. This allowed the relative number of wild
or mutant alleles (hx546) to be
observed directly in each population. In one experiment, Walker et al.
grew the populations under constant feeding conditions and observed no change
in allele frequency over time. Interestingly, this was true over all initial
frequencies of hx546 allele and
suggested that no trade-offs were occurring (see Figure 15a). However, when
the populations were grown under cyclical starvation conditions dramatic
reduction in hx546 mutant allele
was observed (see Figure 15b). This
means that the age-1 genes acted to reduce fertility in simulated real-world
conditions where food was not constantly supplied. Further, they found that
the trade-off between fitness and longevity occurred in young adults (Walker et al.,
2000). The elegance and importance of this study results in
the unity of pleiotropy genetic trade-off theory with implication to real
world versus protected conditions. One of the weaknesses of laboratory
experiments is the large change observed in ageing compared to observations
in the wild (see figure 3).
This study clearly outlines the role of environmental effects on genes which
at first look to be advantages but when in ‘natural’ conditions are clearly
selected against.
Since there was a clear trade-off seen between reproduction and
survival the third concern that was raised by the presence of long-lived
mutants can be effectively answered: 3. The wild type allele for
shortened lifespan was fixed in nature because age-1 mutations experience decreased reproductive capability in
simulated real word environments. |
||
|
|
|
|
|
|
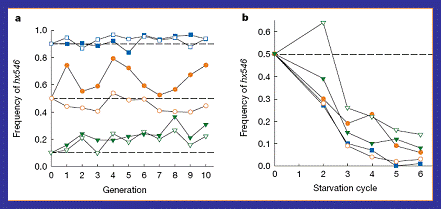
Figure
15:
Results from the study on hx546
which is the mutant allele at the age-1
locus in nematodes. (a) This shows
the lack of change in allele frequency when populations were kept at 20oC
with a constant food source. (b) A
graph showing the rapid decrease in mutant hy546 allele frequency when periods of starvation were added to
the experiment (Walker et al,
2000).
|
Is there always a catch? Known longevity mutants in mice, Drosophila and C. elegans all exhibit some reduction in fitness when placed in natural environments (Voorhies et al., 2006). Since most beneficial mutations will have likely appeared in the past you would expect natural selection to have acted to increase their frequencies in natural populations. For this reason it is suggested that these single gene effects for long life would all have pleiotropic effects and thus would have been selected against early in development. This also implies that the practicality of using such mutants in biomedical research might be limited (Voorhies et al., 2006) |
||
|
|
|
|
|
|
|
A few parting words… Studies
on long-lived mutants do have their place in research on the molecular basis for
ageing. While not contradictory of historical viewpoints, they offer new ways
to test theories and new avenues for learning about complex hormonal pathways
involved in senescence. Furture research will likely use more complex
organisms such as social insects which have natural variation in longevity within
species correlating to behaviour and social status (Keller and
Jemielity, 2006). These studies will
hopefully intergrate genetic mechanisms with real word life-history traits to
better model the ageing process. We
might not be able to find the fountain of youth but research in these fields will
ultimately be used to understand and try to relieve the concequences of time.
|
|
|