|
Classical Evolutionary Theory of Ageing |
||
|
|
|
|
|
|
A challenge to define. Ageing is a concept that, on the surface
at least, seems easy to understand. We think we know what ageing implicitly
means however defining it conclusively has proven challenging. Even published
reviews have asked simply “what is ageing?” and never managed to plainly
define it (Lithgow, 2006).
Ageing research is contemporary and multidisciplinary but it is also rife
with debate over both theory and even the spelling of “ageing” itself (Martin, 2006).
Senescence can be defined as progressive deterioration leading to permanent
loss of function over time (Helfand and Inouye, 2002; Helfand and Rogina, 2003)
and is sometimes used synonymously with ageing. Ageing also refers to the
loss of function as well as the decreasing fertility and increasing mortality
with age (Kirkwood and Austad, 2000). Rate of
ageing can mean the declining contributions to fitness with time (Williams, et al.
2006). It should be noted that ageing,
senescence and lifespan are interpreted differently (or more problematically
interchanged with each other) and this leads to considerable confusion in the
literature and arguments over semantics. Biological markers of ageing vary in appropriateness for
testing theory and thus resulting experimentation has mainly concentrated on
changes in lifespan (Jazwinski, 1998). It is important to note that
longevity (age at death) is not necessarily indicative of ageing rate so
caution must be exercised with this practice (Lithgow, 2006). Other
potential demographic markers which have been used to describe senescence are
survivorship per cohort over time, initial mortality at a certain age, the
rate of increase of age-specific mortality, age specific physiological or
behavioral changes and finally temporal data concerning reproduction and
behavior (Bronikowski and Promislow, 2005). Biological markers that identify
physiological ageing would be extremely valuable for differentiating between
rates of ageing such as temporal gene expression and overall longevity (Helfand and Rogina, 2003). |
|
|
|
|
Figure 2: (a) Vincent Van Gogh(1853 - 1890 ) painting entitled “An Old Woman
From Arles” (b)
A painting of the oldest recorded person in history, Jeanne
Calment who met Van Gogh in her hometown of Arles,
France. She lived until the age of 122 and died in 1997.
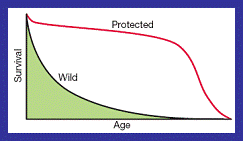
Figure 3: A graph representing the gradual decline in
survival in wild populations with age and the noticeable difference to
individuals who live in protected environments. This result challenges theories
on programmed death and suggested that external environmental factors were
important (Kirkwood and Austad, 2000).
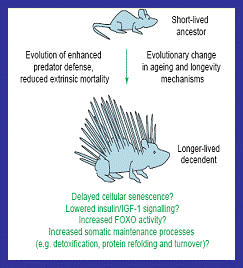
Figure 4: It has been proposed
that more complex defence such as quills, bird wings, or turtle shells led to
increased longevity in those lineages (Partridge and
Gems, 2006).
If no ageing genes should
occur then do long-lived
mutants violate the theory of ageing evolution? Evolutionary theory at one time predicted
that large changes in lifespan could not be caused by single genes but rather
the accumulation of many deleterious mutations over time. Recent studies have
discovered genes which dramatically affect lifespan when mutated. They are
typically involved in the regulation of other pathways but none can cause
immortality. Since the mid-1980’s
there have been studies on long-lived mutants of mostly Drosophila and C. elegans and some will be described in detail in this
website. |
|
|
Can
we say that ageing evolves? There is not a straight
forward answer and depends mostly on your definition of ageing. Complications
arise because ageing is inexorably connected to other biological processes,
namely survival, which clearly undergo evolution and can thus suggest that
ageing has evolved though perhaps not directly. Ageing was considered by some to be a
paradox because although some genes lead to reduction in reproduction and can
undergo evolutionary change, ageing is obviously a maladaptive process (Partridge and
Gems, 2006; amongst others). Most now agree that ageing is
itself not a program (Lithgow, 2006).
The product of ageing is reduced fertility and death which would be selected
strongly against in nature if any so-called “ageing genes” arose. This
viewpoint suggests that ageing cannot evolve. Prior to the advent
of sophisticated genetic research tools, it was hypothesised that ageing was
adaptive for the species. August Weismann first suggested that ageing
occurred because there was natural selection to remove old versions from a
population and free up resources for the young (Partridge and
Gems, 2002; Gavrilov and Gavrilova, 2002). He suggested that organismal death was caused by a genetically programmed
limit in the amount of somatic cellular divisions. All group selection
arguments have come under considerable scrutiny apart from rare exceptions. Natural
selection occurs within a single individual and as such is not able to
maximize the fitness of the entire species with a process that decreases
fitness in the individual itself. Weismann actually changed his opinion on
the matter and stated that instead of being deleterious, old variants are
merely neutral. Testing the theory of programmed death has resulted in
further conformation of its inaccuracies. Unlike predictions, there are
radical differences between the longevities of wild versus protected
individuals. This is true for both laboratory animals and modern humans who
are now able to live to a longer age than ever possible in the wild (see Figures
2&3). This finding is thought to be caused by more benign environments
which decrease the fitness cost of having repair mechanisms that can extend
life (Gavrilov and Gavrilova, 2002). Longevity and
survival must have some genetic component to explain, for example, why
monozygotic twins have more similar life spans than dizygotic
twins (Kirkwood, 2002).
There are also great divergences between the maximum lifespan for various
species and even within species. It is often sited that organisms with
evolved complex behavioural or structural means of escaping predation, such
as flight, tend to have a longer lifespan than their vulnerable ancestors (see
Figure 4). However, the accuracy of this type of claim has come into recent
speculation and most now agree that life-history traits such as flight affect
lifespan in more complex manners (Williams et
al, 2006). A current
area of research concerns this change in ageing rate and the accompanying
changes in other biological processes (Partridge and
Gems, 2006). In a recent paper
documenting the so-called truths of ageing, Lithgow stated that: 1. Genetic programs regulate survival from
the cell cycle level to complex hormonal pathways. 2. Ageing is not
caused by a direct program but is instead the cumulative effect of programs which control survival, fitness
and growth. 3. Life-history
evolution has led to the evolution of different life spans (Lithgow, 2006). So although it is
relatively clear that survival mechanisms are adaptive and selected in nature
based on particular environments, the advent of ageing itself is not
programmed because it does not make evolutionary sense (Lithgow, 2006;
Partridge and
Gems, 2002; amongst others). |
|
|
|
|
|
|
|
Why does ageing occur? After coming to the conclusion that natural
selection cannot select for ageing (the decline in fitness with time) than
this is the logical next question. Thankfully this has a somewhat less
disputed answer based on two main concepts: 1. Since cohort size declines
over time due to illness, predation and accidents, in the wild there are few
old individuals. This coupled with the fact that reproduction typically
decreases with age, means that natural selection does not have as much power
to select against deleterious mutations late in life as it could early on.
Evolution can only act over multiple generations and mutations that occur
post-reproduction are thus irrelevant to natural selection. In wild
populations, it is rare for animals to survive to the period of senescence that
we associate with humans, domesticated and laboratory animals. This further suggests that natural
selection would not be able to act on genes expressed at old age (Kirkwood, 2002). 2. The somatic repair mechanisms
involved in extending longevity counteract mechanisms increasing fitness
early in life (Kirkwood, 2002). The metabolic resources necessary for
survival up to reproduction are more important evolutionarily speaking than
genes encoding maintenance and deterioration repair.
The implications of the first concept of
selection decreasing with age formed the basis for the mutation accumulation and the pleiotropy theories of ageing. The second
concept is the foundation for the disposable soma theory of ageing. Even if one argues that because genes are
effectively hidden from evolution late in life they therefore cannot undergo
natural selection, ageing phenotypes arise from the evolutionary interaction
between survival and genes which ultimately affect how and when ageing
occurs. It is for this reason that the concept of ageing evolving is still
relevant though care should be taken in interpreting any results. There will always be confusion in
terminology and perhaps interpretation because of the multiple layers of
complexity that all have a role in determining ageing in different organisms.
|
|