|
age-1
mutations in C. elegans |
||
|
|
|
|
|
Figure
10 :
An image of two roundworms [Picture from the C.elegans genome project
at the Sanger Institute]. |
Single genes can change longevity Evolutionary theory suggested
that single genes would be unlikely to have large effects on ageing but
rather the accumulation of deleterious mutations over time would lead to
decreased fitness. Caenorhabditis
elegans has long been a model organism for the study of ageing due to its
simple biology and a particular feature of its life called a dauer larva form
(Braeckman and
Vanfleteren, 2006). It is a primarily hermaphroditic
nematode that consists of four larval stages lasting a total of 2.5 days. The
dauer stage is reached when environmental conditions are stressful - lack of food or increased temperature
- and exhibit characteristics of diapause -stress resistance, increased longevity and altered morphology (Braeckman and
Vanfleteren, 2006). Nematodes can stay in the dauer form
for months only to re-emerge once conditions have changed with no sign of
ageing having occurred (Riddle, 1999).
The dauer formation is associated with resistance to oxidative stresses as
well as other environment extremes (Kirkwood and
Austad, 2000). The discovery 20 years ago of a
mutant called age-1 (hx546 allele) which extended the
lifespan of nematodes seemed at first to challenge evolutionary theory in
different ways (Friedman and
Johnson, 1988): 1. The fact that a
single gene could have such a large phenotypic effect meant that some
theoretical ideas about ageing had to be modified. 2. Does the dramatic
change in lifespan due to age-1
mean that ageing is programmed? 3. Why wouldn’t the
long-life variant gene have evolved in natural populations instead of the
wild-type short-lived gene? |
|
|
|
|
|
|
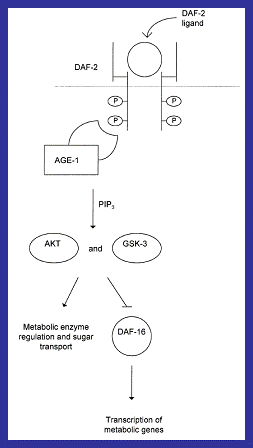
Figure
11: A
simplified representation of how wild AGE-1 decreases longevity by binding to a phosphorylated DAF-2
receptor and activating kinases which down regulate DAF-16 leading to
decreased longevity (Jazwinski,
1998). |
In brief: How age-1 genes are involved in longevity
regulation Grown at 20oC, age-1 mutants and wild-type nematodes do
not vary in “appearance, development rates, activity levels or total
fertility” (Walker et al.,
2000). They do vary in lifespan with the mutants having a
40% (17 - 25 days) increase in mean
lifespan compared to wild (~3 days)
(Friedman and
Johnson, 1988; Kirkwood and
Austad, 2000). This gene was identified as a
phosphatidylinositol-3-OH kinase signal transduction protein belonging to the
insulin/IGF-1 signalling pathway (Jazwinski,
1998). This pathway
is also involved in dauer larval formation as well as stress resistance (see
Figure 11). Further research has found that, like age-1 mutants, organisms with reduced DAF-2 activity live longer
as well as appear younger at old age (Guarente and
Kenyon, 2000). Reduction of function mutations such as age-1 act to increase longevity by
ultimately reducing the level of insulin/IGF-1 signalling which then
activates DAF-16 activity in the nucleus. Once DAF-16 reaches the nucleus
its effectors include stress response genes, antimicrobial and metabolic
genes which found in downstream positions (Braeckman and
Vanfleteren, 2006).
Long-lived mutants would be expected to produce more antioxidant genes
and as a result be more stress resistant (Guarente and
Kenyon, 2000). This was observed in age-1 mutants with the up regulation of catalase activity and
superoxide dismutase. Thus the activities of increased cellular repair
mechanisms reduce ageing in C. elegans
mutants. A response to two of the
above challenges to evolutionary theory are: 1. Though this single gene mutation does
dramatically change ageing rate, its product and associated genetic factors
regulate the affect of numerous genes in the pathway. Thus ageing can still
be considered a polygenic trait 2. The evidence from long-lived mutants does
not change the conclusion that ageing is not a program. That challenge stems
more from misunderstanding of the meaning of a program than what the presence
of long-lived mutants means for evolution theory. The presence of the mutants
merely reinforces the notion that ageing is affected by genetic variation not
that this genetic variation directly programs an ageing phenotype. (Partridge and
Gems, 2006). Over the last two decades
considerable research has further explored the intricacies of this pathway (see figure 14).
The regulation in longevity by the insulin/ IGF-1 pathway is observed in both
Drosophila and mice which suggest
these long-lived mutant genes might have more widespread implications for
ageing research (Braeckman and
Vanfleteren, 2006). |
|
|
|
|
|
|
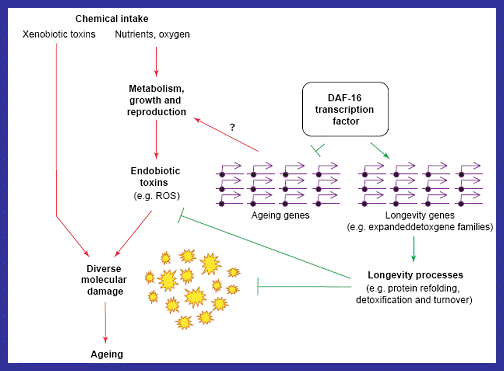
Figure 12: A diagram illustrating the molecular basis for ageing and longevity in IGF-1 signalling. The transcription factor DAF-16 stimulates the expression of genes encoding products that increase the longevity of cells by reducing damage caused by extrinsic factors (Partridge and Gems, 2006). |
||
|
|
|
|
|
Germ line vs. Somatic signalling Reproduction system signals also regulate lifespan with the DAF-2/DAF-16 pathway. In a study by Hsin and Keyon, animals lacking daf gene functions underwent laser surgical ablation (Riddle, 1999). Ablation of germ line cells resulted in an increase in lifespan of 60%, while ablation of the somatic gonad did not affect lifespan (Guarente and Kenyon, 2000). The results of this study suggested that normal somatic gonad signals promote longevity by down regulating DAF-2 which acts to shorten life. Germ line signals thus should decrease longevity by down regulation of DAF-16 which acts to extends life (Braeckman and Vanfleteren, 2006). Figure 13 : Two representations of the trade-off between germ line and somatic-gonad signalling ultimately regulating the longevity of C.elegans and occurring as predicted by the disposable soma theory ((a) Riddle, 1999 (b) Guarente and Kenyon, 2000) |
|
|
 (a)
(a)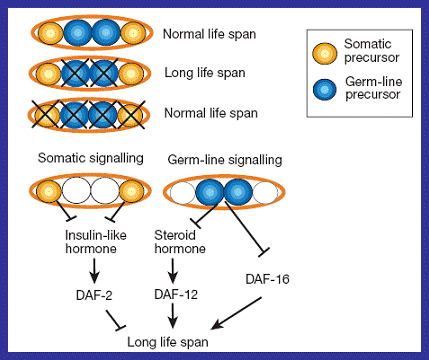 (b)
(b)